This June I participated in an oceanographic research cruise to Barbados. One of the goals of this expedition was to collect planktonic larvae of benthic marine invertebrates using a MOCNESS device. Among other things, we found larvae of inarticulate brachiopods (phylum Brachiopoda; class Inarticulata) pictured here. Brachiopods resemble bivalve molluscs (e.g. clams and mussels), but have dorsal and ventral valves rather than left and right. The valves are held together with muscles in inarticulate brachiopods, while those of articulate brachiopods are hinged. Brachiopods, as a phylum, are also characterized by the lophophore (a crown of tentacles surrounding the mouth). We collected two different kinds of inarticulate brachiopod larvae.

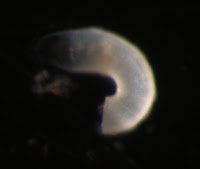
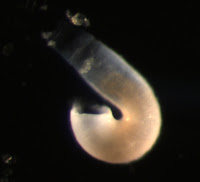
Larvae of the first kind were collected from relatively shallow depths (900-0 m). In this image, one can see the large circular larval shell valves, as well as the embryonic shell (the half circle at the posterior portion of the larval shell, tinged blue). The lophophoral tentacles are retracted in the first image, but extended in the second.

These tentacles form progressively during the larval life from short tentacle buds on either side of a longer unpaired anterior median tentacle (at about 12 o’clock on the second image). Consequently, the number of tentacles reflects larval age. This larva has 10 pairs of tentacles. In some species, a lophophore with this many tentacles indicates larval competence for settlement (Pennington and Stricker, 2002). One can also see that the tentacles of the lophophore are ciliated. Cilia are used for larval feeding and swimming (Rudwick 1970). This particular type of larva is produced by members of the superfamily Lingulacea which contains only two extant genera: Lingula and Glottidia (Pennington and Stricker 2002).
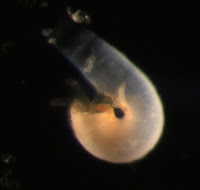
 Larvae of another kind were collected from deeper waters (1600-900 m). Characteristically, they lack an embryonic shell, and have a pair of larval chaetae. Presence of chaetae in this larva suggests that it belongs to the inarticulate superfamily Discinacea (Pennington and Stricker 2002). Chaetae are usually thought of as a feature unique to annelid worms, but some brachiopod larvae and adults also have chaetae, which are similar structurally and developmentally to those of annelids.
Larvae of another kind were collected from deeper waters (1600-900 m). Characteristically, they lack an embryonic shell, and have a pair of larval chaetae. Presence of chaetae in this larva suggests that it belongs to the inarticulate superfamily Discinacea (Pennington and Stricker 2002). Chaetae are usually thought of as a feature unique to annelid worms, but some brachiopod larvae and adults also have chaetae, which are similar structurally and developmentally to those of annelids.
Pennington, J T and S A Stricker. 2002. Phylum Brachiopoda. In: Atlas of Marine Invertebrate Larvae. Edited by Craig M Young. Academic Press.
Rudwick, M J S. 1970. Living and Fossil Brachiopods. Hutchinson & Co, London.
















{kind=link}
{kind=link}